







Sfarbenie hláv a štruktúra peria u amadín Gouldovej, Gouldian finches
Sfarbenie hlavy u amadín Gouldovej
Typy sfarbenia hlavy
U amadín Gouldovej v prírode žijú 3 farebné variácie s prírodným sfarbením tela. Sú to exempláre s červenou hlavou, s čiernou hlavou a so žltou (oranžovou) hlavou.
Otázkou je, ktorá z nich je najpravdepodobnejšia nominátna forma, alebo naozaj máme dočinenia s tromi rozdielnymi druhmi alebo poddruhmi ?
Červenohlavé vtáky sa považujú za nominátnu formu žijúcu vo voľnej prírode. Zvyšné dve formy sú pravé mutácie, ktoré vznikli v prírode. Nič nevylučuje, že by mutačný jedinec nemohol prežiť v prírode, avšak väčšinu času sú tieto vtáky tak jasne viditeľné alebo bezmocné, že sú ľahkým terčom pre ich prirodzených nepriateľov. Preto mutačným vtákom hrozí vyhynutie. Skutočnosť, že sa zmutované amadiny Gouldovej nachádzajú v prírode, môže byť spôsobená len „malou“ mutáciou farby hlavy. Vtáky s týmto „malým rozdielom“ sú pre predátorov sotva rozpoznateľné medzi zvyškom populácie. Pri príkladoch s párením si ukážeme, že nejde o medzidruhové kríženie.


Behaviorálne a genetické otázky
Nedávny výskum Pryke-a a Griffith-a popísal rozdiely v správaní sa červenohlavých a čiernohlavých amadín Gouldovej (Pryke a Griffith, 2009).
V prírode čiernohlavé vtáky prevyšujú červenohlavé vtáky v pomere 3:1. Predpokladá sa, že v dávnej minulosti boli červenohlavé a čiernohlavé amadiny Gouldovej geograficky oddelené populácie a existovali ako dva rozdielne „poddruhy“- v zmysle jeden druh teritoriálne žijúci v oddelených spoločenstvách. Až neskoršie prišli znovu do kontaktu a začali koexistovať ako jedna populácia. Toto pôvodné geografické oddelenie môže byť vysvetlením, prečo sú prítomné také výrazné rozdiely v správaní amadín Gouldovej u jednotlivých typov sfarbenia hláv.
Samčeky aj samičky s červeným sfarbením hlavy sú agresívnejšie (priebojnejšie) ako čiernohlavé vtáky. Súťaživejšiu povahu červenohlavých vtákov možno vysvetliť obmedzeným prístupom k hniezdnym dutinám, vode a potravinovým zdrojom v areáli, ktorý obývali počas ich geografického odlúčenia.
„Pasívnu“- menej priebojnú povahu čiernohlavých amadín Gouldovej je možné pripísať prostrediu bohatému na potravu, takže vtáky nemuseli o potravu súperiť, a preto v ich povahe nie je toľko geneticky zakódovanej súťaživosti.
Predchádzajúci výskum poukázal na vyššiu úmrtnosť samičiek v porovnaní so samčekmi vtedy, keď dochádza ku páreniu jedincov s rôznym zafarbením hlavy. Pryke a Griffith zistili, že pri párení červenohlavých s čiernohlavými prežije menej potomkov v porovnaní s párom s rovnakou farbou hlavy. Toto zistenie nebolo pozorované v chovoch na vtákoch, ktoré pochádzali z voľnej prírody. I keď išlo o vtáky z prírody, čiernohlavé vtáky si často vyberali červenohlavých partnerov/partnerky, hoci mali možnosť vybrať si partnera s takou istou farbou hlavy, ako majú oni. Párenie medzi čierno- a červenohlavými vtákmi môže byť jedným z dôvodov poklesu počtu kusov voľne žijúcej populácie. Za normálnych podmienok amadiny Gouldovej preferujú párenie s vtákmi tej istej farby hlavy. Páry s čiernou farbou hlavy sú schopné za prítomnosti genetických dispozícií mať červenohlavé potomstvo a červenohlavé páry sú schopné mať potomkov všetkých troch farieb. Hoci červenohlavé vtáky majú obrovskú predispozičnú výhodu genetickej dominancie nad čiernohlavými (t. j. obsadzujú najlepšie hniezdne dutiny, pijú prvé z napájadiel atď.), je ich v populácii menej vďaka ich sociálnej agresivite, vyššej náchylnosti na stres a vyššej hladine pohlavných hormónov... Tieto vlastnosti sú predispozíciou k zdravotným problémom, ako je roztočová infekcia vzduchových vakov, a vďaka vyššej náchylnosti na stres nie sú schopné udržať si normálny stupeň imunity. V čase, keď počty stavu voľne žijúcej populácie vtákov dramaticky klesali ich odchytom, sa pravdepodobne väčšia časť samičiek z nedostatku možnosti výberu partnera spárila so samčekmi (t.j. červeno- a čiernohlavé párenie). Vyšší počet týchto rozdielnych dvojíc mohol mať negatívny vplyv na schopnosť miestnej populácie rozrastať sa vzhľadom na zníženú pravdepodobnosť prežitia mláďat a zvýšenému počtu samčekov medzi prežívajúcim potomstvom (Pryke a Griffith, 2009). Konečné výsledky zaznamenali vyšší úhyn samičiek z takéhoto párenia (Pryke a Griffith, 2009).
Poznámka 1.: Tu by som chcel upozorniť na fakt, že teória o genetickej rozdielnosti medzi červenohlavými a čiernohlavými vtákmi nebola v zajatí preukázaná. Aspoň nie do takej miery, aby bola aplikovateľná v chovoch. Myslím na skúsenosti chovateľov. Ani ja osobne takúto skúsenosť nemám. Štúdium voľne žijúcej populácie sa zameriava na vtáky, ktoré žili geograficky oddelené po stovky generácií. Zhodou okolností jedna z nich mala červenú hlavu a ďalšia čiernu. Samotná farba hlavy však podľa mňa nehrá úlohu, dôležité je, že červenohlavé vtáky žili v nehostinnom prostredí, ktoré ich nútilo k súťaživosti a teda aj k vyššiemu stresu. Čiernohlavé vtáky však žili v pohostinnejšom prostredí a preto sa u nich súťaživosť, ani predpoklady pre stres, tak nevyvinuli. Ak bolo odlúčenie jednotlivých populácií dostatočne dlhé, tieto vlastnosti sa zapísali aj do genetických predispozícií a tu môžeme hľadať korene „nezlučiteľnosti“ červeno- a čiernohlavých vtákov. Je vysoký predpoklad, že voľne žijúca populácia si tieto rozdielnosti zapísané v génoch stále uchováva. V chovoch boli však vtáky už pomerne dlhú dobru párené bez ohľadu na farbu hlavy a dá sa teda predpokladať, že génová diverzita sa aspoň čiastočne zotrela. Podľa môjho názoru sa počas domestikácie genetická výbava nesúca vlastnosti popísané na divo žijúcej populácii už dostatočne premiešala. Stále však ostáva priestor na výskum ako v prírode, tak i v chovoch a, samozrejme, vždy je otvorená otázka na diskusiu.
Poznámka 2.: V poslednej dobe niektorí biológovia venujúci sa evolučnej biológii prišli s názorom, že od nominátnej červenohlavej amadiny Gouldovej sa odčlenil poddruh, a to čiernohlavé amadiny Gouldovej. Podľa nich je súčasný stav akýmsi medzistupňom na vytvorenie dvoch nezávislých druhov. Na to je však potrebné, aby populácie boli dosť silné a aby žili geograficky oddelene, i keď sú známe teórie, že nový druh môže vzniknúť v rovnakom teritóriu ako pôvodný, pokiaľ sa špecializoval. Ako argument na podporu poddruhov sa uvádza zvýšená úmrtnosť mláďat pri párení červenohlavých a čiernohlavých amadín Gouldovej (vychádzajú z medzidruhového kríženia). Objavili sa tiež teórie o tom, že červenohlavá samička si vyberá červenohlavého samčeka, a s týmto má početnejšiu znášku a viac oplodnených vajíčok, a v závislosti na farbe hlavy partnera dokáže regulovať pohlavie vyliahnutého potomstva. V každom prípade si myslím, že tieto teórie sú nové, zatiaľ neoverené, a čas ukáže, či aj pravdivé. Pomôcť snáď môže aj molekulárna biológia, ktorá by vec rozsúdila, i keď nie každý súd je vždy definitívny.
Párenie v chovoch
I keď beriem do úvahy zahraničné štúdie venujúce sa divožijúcej populácii, v chove som nezaznamenal žiadne problémy pri zostavovaní párov s rôznym sfarbením hlavy. Úhyn nebol väčší ani menší, ak mal rodičovský pár rovnaké, či rozdielne sfarbenie hláv. Nevidím dôvod ani na to, aby sa chovali „čisté“ línie sfarbenia hlavy. Nezaznamenal som žiaden problém, aj keď sa do chovu počas niekoľkých generácií zaraďovali páry s rovnakým sfarbením hlavy, či už s čisto červenými, čiernymi, či žltými hlavami. Nemám ani takú skúsenosť, že by párenie žltohlavých vtákov počas viac generácií viedlo k degradácii (poruche) operenia. Je však dôležité, aby chovateľ do chovu zaraďoval vtáky prísne selektívne, teda kondične a fenotypovo dostatočne silné. Čo sa týka párenia vtákov s rôznym sfarbením hlavy, tak je skôr dôležité, k akému potomstvu sa chce chovateľ dopracovať a podľa toho by mal rodičovský pár zostaviť.
Druh peria na hlave
Máme dočinenia s dvomi typmi peria. Perie červenohlavých a žltohlavých vtákov je iné ako perie čiernohlavých.
Žltohlavé a červenohlavé: Sú rozdiely aj v štruktúre pierok samičiek a v štruktúre pierok samčekov u červenohlavých a u žltohlavých vtákov:
Samčie pierka majú dlhé vetvičkami bez lúčov s háčikmi na konci. Odhora nadol: je červená časť, potom hnedá časť, nasleduje malá čierna časť. Spodná časť pera je sivo- biela.
Samičie vetvičky bez lúčov s háčikmi na konci sú omnoho kratšie. Odhora nadol: červená časť, potom hnedá časť, potom variabilne relatívne sýta čierna časť a spodok pera je ako u samčekov. Samičky majú typicky tmavšie sfarbenú červenú a často aj žltú hlavu práve vďaka tejto štruktúre a vďaka rozdielom v proporciách medzi jednotlivými farbami v sfarbení pera.
Tieto pierka sú ornamentálneho typu a obsahujú červený kantaxantín a žltý luteín - (karoténove prírodná farba). Ak je produkcia kantaxantínu na hlave prerušená, výsledkom je vytvorenie žltého luteínu.

Čiernohlavé vtáky : operenie hlavy u čiernohlavých samičiek a samčekov je takmer identické. Celá zástavica je pokrytá malými háčikmi. Pierka sú úplne vyplnené čiernym eumelanínom. Inými slovami, v porovnaní s pierkami červenohlavých vtákov tieto pierka sú menej vyvinuté a chýba im „špička“, kde je zvyčajne umiestnené červený alebo žltý pigment.

{kind=link}
V perí amadín Gouldovej nachádzame 4 základné pigmenty:
Melaníny:
čierny ( eumelanín). Najviac sa ho nachádza v čiernom zafarbení hlavy

hnedo - červený (pheomelanín). Najviac sa ho nachádza v perí na prsiach, kde spôsobuje vytváranie modro-fialového zafarbenia. Je teda zodpovedný za tvorbu štrukturálnej farby podobne ako eumelanín. Prítomný je však aj v ostatnom operení tela. Ilustrujem to na fotkách hlavy žltohlavých vtákov, kde je jasne vidieť, že zelený fialovoprsý a zelený bieloprsý vták má odlišné sfarbenie peria hlavy. To znamená, že jeho neprítomnosť ovplyvňuje celkovú intenzitu sfarbenia. Bieloprsé vtáky majú jasnejšie farby.
Karotenoidy:
žltý (luteín epoxid – alfa xanthofyl). Na fotke žltohlavého vtáka je ilustrované nielen rozloženie luteínu, ale v komôrkovej vrstve je prítomný tiež pheomelanín
červený (canthaxanthin (kantaxantin) – beta karotenoid). Jeho intenzitu je vidieť na fotkách červenohlavých vtákov. Podobne ako u žltohlavých je prítomný aj pheomelanín, čo má za následok, že bieloprsé vtáky majú pre neprítomnosť pheomelanínu jasnejšie sfarbenie hlavy.

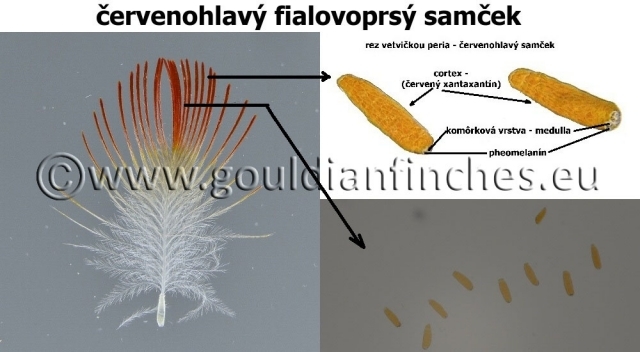
Ako je vidieť na priloženej fotografii, intenzita sfarbenia v strednej časti pera „1“ je ovplyvnená pheomelanínom. U fialovoprsých vtákoch je prítomná, u bieloprsých prakticky chýba. Prítomnosť hnedého sfarbenia nie je závislá od toho, či je vták zelený alebo 2 faktorový žltý. Časť označená ako „2“ je zase závislá od mutácie zeleného sfarbenia. U zelených vtákoch je prítomný čierny eumelanín, u dvojfaktorových žltých prakticky chýba, farba pŕs túto časť operenia neovplyvnila.
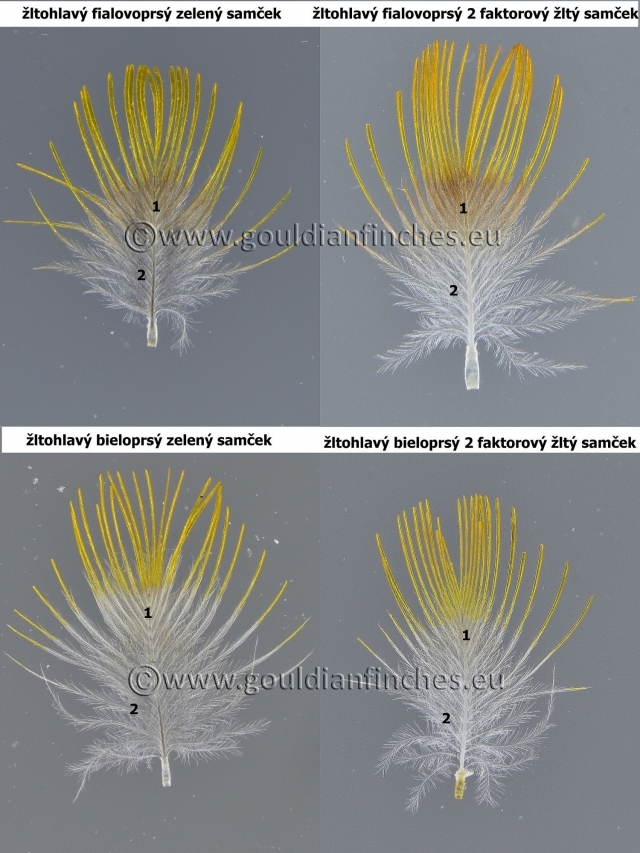
U amadín Gouldovej bolo vyšľachtených veľa farebných mutácií. Mutácia sfarbenia tela ovplyvňuje aj fenotyp sfarbenia hlavy.
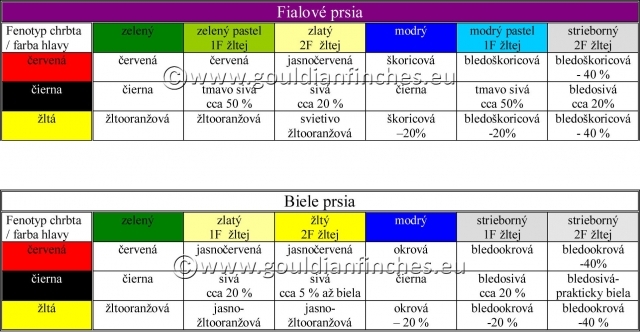
Dedičnosť farieb u mutácií je však rovnaká ako u prírodne sfarbených vtákov. Preto hovoríme o červenohlavom zelenom, modrom, či striebornom, i keď fenotypovo je sfarbenie odlišné. Analogicky to platí aj u ostatných farieb hlavy. V priložených tabuľkách popisujem zhruba odtiene, aké sú pri jednotlivých mutáciách. Na fotografiách vidíte niektoré farebné mutácie a rozdielnosti v sfarbení hlavy. Dôležité je všímať si najmä celkovú štruktúru pierok, oblasť s pheomelanínom (pri fialovoprsých výraznejšia) a oblasť spodných bočných vetvičiek s eumalenínom v závislosti od mutácie. U zelenej a modrej je pigmentácia zachovaná, u žltej je potlačená.
{kind=link}
Skratky a označenia
Opustím moje zvyčajné značenie, teda aj to, ktoré je použité v genetickej kalkulačke a skúsim dedičnosť vysvetliť na inom, mierne odlišnom princípe. Budeme predpokladať, že červená farba je dominantná a viazaná na pohlavie, čierna farba je recesívna a viazaná na pohlavie. V tomto prípade teda čiernu považujeme za recesívnu alelu dominantnej alely pre červenú. Mechanizmus vzniku pigmentov som opísal v inej kapitole, tak len v skratke. Ak v procese tvorby červeného farbiva – kantaxantínu - nastane zmena, príde k vytvoreniu žltého luteínu. Gén, ktorý spôsobí prerušenie tvorby červeného farbiva je na autozomálnom chromozóme, nie je teda viazaný na pohlavie a dedí sa voľne. Tento gén je modifikátor, ktorý je buď aktívny, a tak znemožní vytvorenie červeného farbiva, alebo je neaktívny a príde k vytvoreniu červeného farbiva.
RR – červená farba hlavy
rr - čierna farba hlavy
ll – aktívny modifikátor na zastavenie tvorby červenej (tvorí sa žltá)
ii - neaktívny modifikátor na zastavenie tvorby červenej (tvorí sa červená)
P- parietálna - rodičovská generácia
F1 - 1. Generácia potomkov P
F2 - 2. Generácia potomkov
G- gaméty (pohlavné bunky)
♂ Samci homozogotní

1.Červenohlaví RR (Red) - majú červenú špičku zobáka: farba je viazaná na pohlavie a dominantná.
2.Čiernohlaví rr (black) - majú červenú špičku zobáka: farba je viazaná na pohlavie a recesívna. Čierne sfarbenie hlavy je neprítomnosť červenej farby, a tak sa zdá, že je to recesívna alela pre červenú. Keďže je táto farba viazaná na pohlavie a recesívna, a samčeky majú dva pohlavné XX chromozómy, môžu na túto farbu štepiť.
3.Žltohlaví II (orange) - majú žltú špičku zobáka: v genotype sa prejaví len v dvojfaktorovom stave, teda ak sú aktívne obe alely modifikátora pre zastavenie tvorby červenej.
Modifikátor pre tvorbu žltej farby je umiestený na autozomálnom chromozóme, dedí sa teda nezávisle od pohlavia. Žltá musí byť prítomná v DF, aby sa prejavila vo fenotype, je recesívna, výnimku tvorí recesívna epistáza. Aby sa žltá farba prejavila vo fenotype hlavy, musia byť na pohlavných chromozómoch XX umiestnené gény pre červenú farbu hlavy. U žltej farby udávam, že je recesívna, ale len v prenesenom slova zmysle. Keď je v heterozygotnom - štepiteľnom stave, tak sa voči skúmanému génu správa podobne ako rôzne alely jedného páru - neprejaví sa vo fenotype.
Ii: v genotype je prítomný jeden faktor zmutovanej žltej (štepiteľná na žltú).
ii: nie je aktívny modifikátor pre tvorbu žltej farby
4.Čiernohlavé YTB vtáky so žltou špičkou zobáka. rr;II (YTB). Sú geneticky dvojfaktorové žltohlavé vtáky, s fenotypom čiernej farby na hlave, ktorá znemožní expresiu žltej. Ide o potlačenie žltého (luteínu) čiernym (eumelanínom). Modifikátor pre žltú je teda aktívny, avšak na pohlavných chromozómoch sa nachádzajú alely pre čierne sfarbenie.
♀Samičky hemizygotné a homozygotné

1.Červenohlavé RY (Red) - majú červenú špičku zobáka: farba je viazaná na pohlavie a dominantná. Keďže sú samičky pre túto farbu hemizygotné, postačuje jedna alela na prejav znaku.
2.Čiernohlavé RY (black) - majú červenú špičku zobáka: farba je viazaná na pohlavie a recesívna voči červenej. Čierne sfarbenie hlavy je neprítomnosť červenej farby, a tak sa zdá, že je to recesívna alela pre červenú. Označujeme ju ako neprítomnosť R červenej. Keďže je táto farba viazaná na pohlavie a recesívna a samičky majú len jeden pohlavný X chromozóm, nemôžu na túto farbu štepiť.
3.Žltohlavé II (orange) - majú žltú špičku zobáka: v genotype sa prejaví len v dvojfaktorovom stave, teda ak sú aktívne obe alely modifikátora pre zastavenie tvorby červenej.
Modifikátor pre tvorbu žltej farby je umiestený na autozomálnom chromozóme, dedí sa teda nezávisle od pohlavia. Žltá musí byť prítomná v DF, aby sa prejavila vo fenotype, je recesívna, výnimku tvorí recesívna epistáza. Aby sa žltá farba prejavila vo fenotype hlavy, musia byť na pohlavných chromozómoch XX umiestnené gény pre červenú farbu hlavy. U žltej farby udávam, že je recesívna, ale len v prenesenom slova zmysle. Keď je v heterozygotnom - štepiteľnom stave, tak sa voči skúmanému génu správa podobne ako rôzne alely jedného páru - neprejaví sa vo fenotype.
Ii: v genotype je prítomný jeden faktor zmutovanej žltej (štepiteľná na žltú).
ii: nie je aktívny modifikátor pre tvorbu žltej farby
4.Čiernohlavé YTB vtáky so žltou špičkou zobáka. rY; II (YTB). Sú geneticky dvojfaktorové žltohlavé vtáky, s fenotypom čiernej farby na hlave, ktorá znemožní expresiu žltej. Ide o potlačenie žltého (luteínu) čiernym (eumelanínom). Modifikátor pre žltú je teda aktívny, avšak na pohlavnom X chromozóme sa nachádza alela pre čierne sfarbenie.
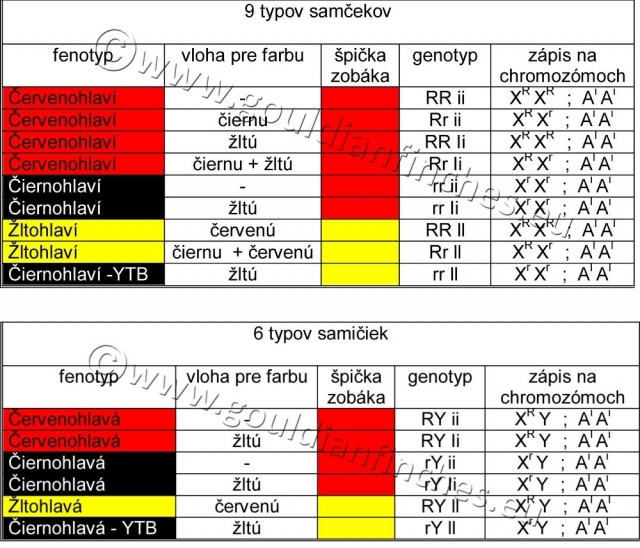
Z týchto definícií dostávame 9 typov samčekov a 6 typov samičiek (homo a heterozygotných jedincov).
Príklady párenia
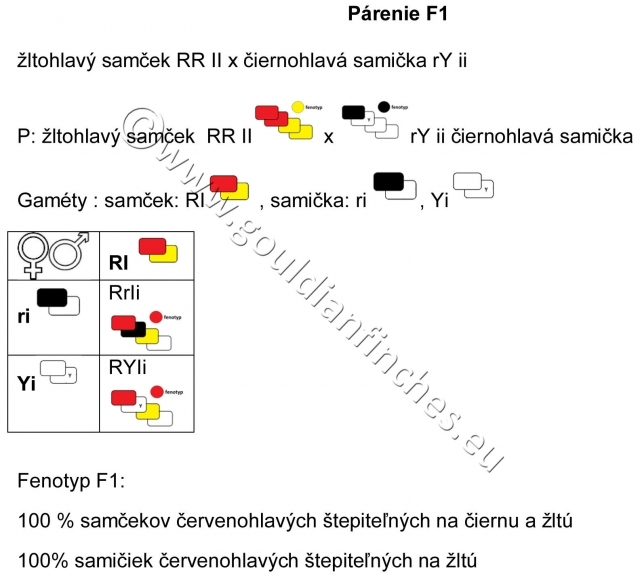
Ak by tieto formy boli rozdielne druhy, potom by sme mali získať potomstvo krížencov z párenia medzi rozdielnymi druhmi. Avšak, tu nejde o takýto prípad, pretože ak spárime čiernohlavého samčeka s červenohlavou samičkou, potom získame čiernohlavé samičky a červenohlavých samčekov štepených na čiernu farbu hlavy.
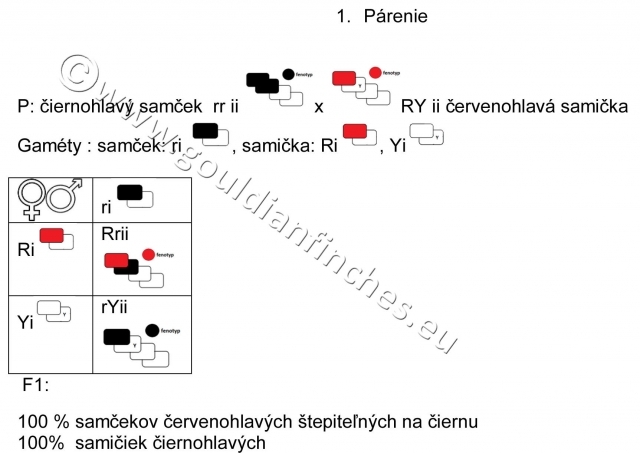
Ak spárime červenohlavého samčeka s čiernohlavou samičkou, tak všetky mláďatá budú červenohlavé, ale samčeky budú štepiť na čiernu farbu hlavy. Je veľmi dôležité to vedieť, pretože to ukazuje na to, že gén zodpovedný za čiernu farbu hlavy a gén pre červenú farbu operenia je umiestnený na pohlavnom chromozóme, a preto je viazaný na pohlavie. Gén pre žltú farbu hlavy sa dedí ako autozomálny znak, a preto je umiestnený v autosomálnom chromozóme, teda nie je viazaný na pohlavie.
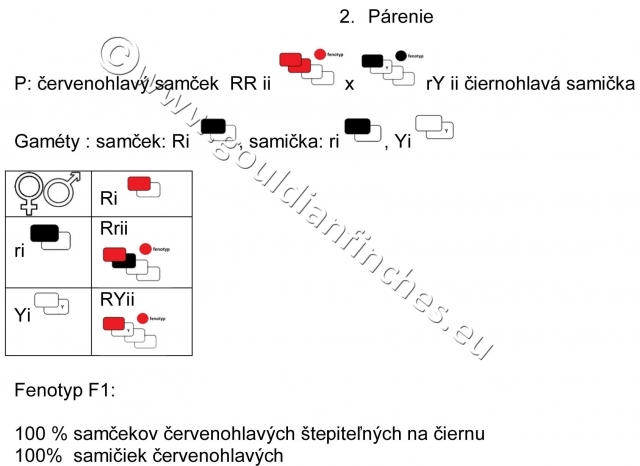
Ak párime žltohlavého samčeka s čiernohlavou samičkou, tak celé potomstvo bude červenohlavé. Všetky samčeky budú štepiteľné na čiernu farbu hlavy a oranžovú farbu hlavy a samičky budú štepiteľné na žltú farbu hlavy.
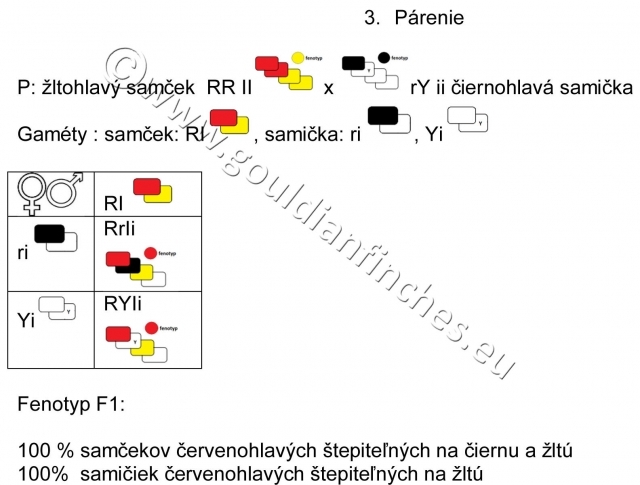
Spárime čiernohlavého samčeka štepiteľného na žltú a žltohlavú samičku. Obaja rodičia nesú vo fenotype len recesívne sfarbenie , teda čierne a žlté. Potomkovia však budú mať vo fenotype aj dominantnú červenú farbu.
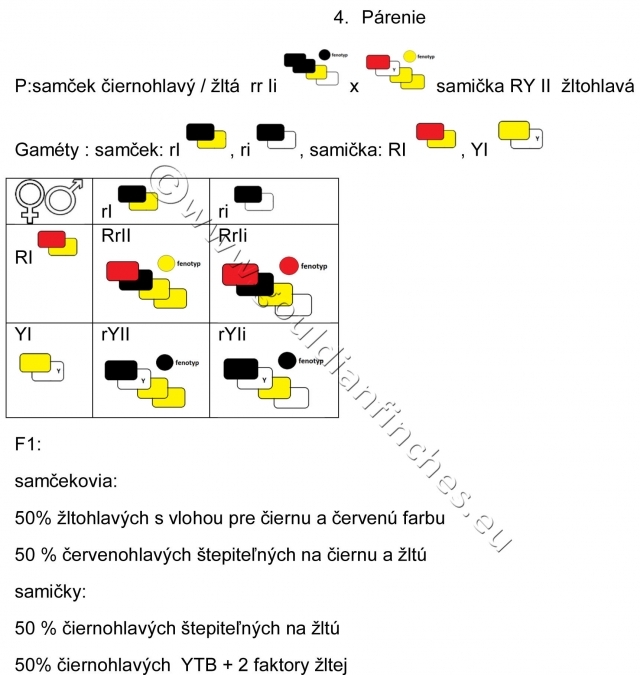
Spárenie čiernohlavého samčeka so žltohlavou samičkou ukazuje na génovú interakciu. Výsledkom bude potomstvo, ktoré nesie farbu rodičov, ako i dominantnú červenú farbu, ktorú rodičia vo fenotype nemali.
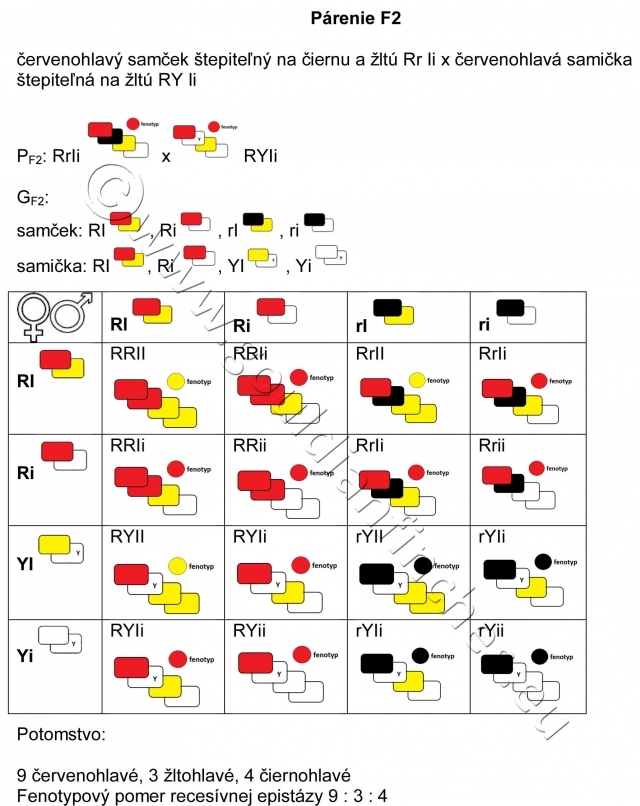
Pri tomto párení vzniká zdanlivý paradox, keď akoby neplatili zákony genetiky. Teda od dvoch rodičov, ktorí nesú recesívne znaky vo fenotype dostaneme potomstvo s dominantným sfarbením hlavy, teda červenohlavé. Poučky z genetiky nám v zásade hovoria, že vo fenotype sa presadí dominantný znak – farba, a že živočích, teda aj vtáky, nemôžu byť nositeľmi vlastnosti pre dominantný znak - farbu, pokiaľ sa táto neprejaví vo fenotype. Príklad génovej interakcie – recesívnej epistázy - však hovorí aj o tom, že je možné, aby fenotyp niesol recesívny znak a pritom v genotype bude zakódovaná vlastnosť (štepiteľnosť) na znak dominantný. S takýmto príkladom sa môžeme stretnúť napríklad pri dedičnosti hláv u amadín Gouldovej.
Definícia recesívnej epistázy: ak sú prítomné 2 recesívne alely v homozygotnej konštitúcii, môžu potlačiť fenotypový prejav dominantnej alely na inom lokuse v homozygotnej alebo heterozygotnej konštitúcii.
To teda znamená, že sa vo fenotype – vzhľade potomstva môže objaviť dominantná farba, ktorá nebola viditeľná vo fenotype rodičov. Inak povedané, vtáky s fenotypom recesívnej farby môžu niesť vlohu (byť štepiteľné) na dominantnú farbu.
To je prípad aj párenia číslo 4. U samčekov 2 faktory žltej potlačili vo fenotype červenú, obdobne je to u samičky, i keď tá je pre červenú hemizygotná, má teda len jednu alelu pre daný znak.
Dôkaz cez štiepne pomery, že ide o recesívnu epistázu
Fenotypový pomer recesívnej epistázy 9:3:4,
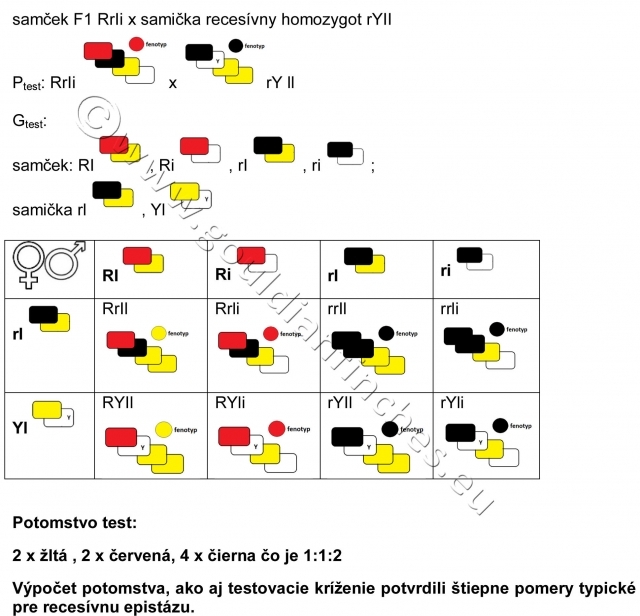
Fenotypový pomer recesívnej epistázy pri testovacom krížení: 1:1:2.
P- parietálna (rodičovská generácia)
G – gaméty (pohlavné bunky)
F1 – prvá filiálna generácia
F2 – druhá filiálna generácia rodičov z F1
Testovacie kríženie (kríženie F1 generácie s recesívnym homozygotom)
Niekoľko príkladov sfarbenia peria hlavy u vybraných mutácií v ďalšej kapitole
Odoberanie vajíčok a ich následná inkubácia
02.11.2016 - Kniha o chove amadín Gouldovej.
http://gfinches2.com/
http://www.gouldamadine.de/
http://www.novaexota.eu/
http://www.epapousek.cz/

stránok www.gouldianfinchces.eu
Kontakt: